
The Book of Mormon and the Origin of Native Americans from a Maternally Inherited DNA Standpoint
Ugo A. Perego
Ugo A. Perego, “The Book of Mormon and the Origin of Native Americans from a Maternally Inherited DNA Standpoint,” in No Weapon Shall Prosper: New Light on Sensitive Issues, ed. Robert L. Millet (Provo, UT: Religious Studies Center, Brigham Young University, 2011), 171–217.
The consensus among scholars is that America’s double continent was the last of the world’s landmasses to be colonized by modern humans, probably by crossing the massive Beringian land bridge that once connected Northern Asia to Alaska. However, the Book of Mormon tells that through divine guidance, relatively small groups of Old World migrants were led to the Western Hemisphere from time to time. Critics argue that since the DNA of Native Americans shares Asian rather than Middle Eastern affinities, the Book of Mormon is fictional and not historical in nature.
Where did Native Americans come from? When did they arrive in the Western Hemisphere? Which route(s) did they follow? How many colonization events were there? These and other fascinating questions have been at the center of the debate among scholars from different disciplines since the rediscovery of the New World by Europeans more than five hundred years ago. Archaeologists, linguists, anthropologists, and geneticists are still investigating the processes that took place through the millennia that led to the peopling of America’s double continent. The considerable number of scholarly papers that have been published on DNA and Amerindians is a demonstration that “despite the 80-year history of genetic studies in the Americas, the real work is now [only] beginning to fully elucidate the genetic history of [the] two continents.” [1]
At first, Europeans believed that the New World inhabitants were somehow connected with the biblical account of the lost ten tribes (see 2 Kings 17:6), leading them to look for cultural and linguistic similarities between contemporary Jews and Native Americans. [2] The evidence amassed to this point indicates that although sporadic pre-Columbian contacts with the Old World cannot be completely ruled out, [3] the majority of Native Americans share a genetic affinity with Asian populations. [4]
The notion that some, or all, American Indians are of Hebrew descent is still popular among Latter-day Saints. The Book of Mormon tells of three relatively small parties (the Jaredites, Lehites, and Mulekites) that left their native homeland in the Old World at different times and through divine guidance traveled to a new promised land, on the American continent. The Book of Mormon contains only marginal information about the demographic dynamics and the geography of the land occupied by the people it describes. Instead, the volume claims to be an abridgment of thousands of years of mostly spiritual and religious history and not a full account of the people. For example, the text does not give direct information about whether other populations were already established in the land at the time of the migrants’ arrival. This lack of information leaves many open questions that have profound implications for the genetic characteristics that we would expect to find in present-day Native American populations. The extent to which these Old World groups expanded and colonized their new habitat, the level of admixture they may have experienced with local indigenous populations (if any were present), and the locations of their settlements would all influence the genetic landscape we would observe in Native Americans today. Furthermore, it is implausible that ancient record keepers would have had a comprehensive knowledge of all the goings-on of the entire vast landmass of the Americas, considering that the distance from northern Canada to southern Patagonia is about 8,700 miles, a greater distance than that from Portugal to Japan! Despite these many complex factors, since the publication of the Book of Mormon in 1830, Mormons and non-Mormons alike have resorted to speculation in an attempt to fill in the historical and geographical details that are either completely missing or only briefly alluded to in the Book of Mormon text. [5]
Even in light of statements by individual Latter-day Saint Church leaders and scholars on this topic through the years, the Church advocates no official position on the subjects of Book of Mormon geography and the origins of Amerindian populations. [6] Together with all other members, Latter-day Saint Church leaders are entitled to their own opinions and reasoning on this subject, as demonstrated by “pre-DNA” comments such as that of President Anthony W. Ivins, a member of the First Presidency, at the April 1929 general conference: The Book of Mormon “does not tell us that there was no one here before” the Book of Mormon peoples. “It does not tell us that people did not come after.” [7] Others have expressed similar opinions more recently. [8]
Over the past decade, critics of the Book of Mormon have promoted the idea that since the majority of Amerindian DNA lineages are closely related to Asian populations, and since no perfect genetic affinity to the Middle East has been found, it must be concluded that the Book of Mormon account is fictional. This argument is sometimes bolstered in part by a common sentiment among Latter-day Saints generally that all Native Americans are descendants of the Old World migrants described in the Book of Mormon text, particularly Lehi’s colony. To contend with these arguments, some Mormons dismiss DNA studies as being unreliable for reconstructing history, while others are quick to embrace any news of possible Middle Eastern DNA in the Americas as conclusive proof that the migrations to America described in the Book of Mormon are real.
In this article, I will provide an updated review on the properties of mitochondrial DNA (mtDNA) and explain how these pertain to the study of ancient population expansions, specifically focusing on the origin of Native Americans. This topic is especially relevant to the current debate on the applicability of DNA evidence to the question of Book of Mormon historicity, as such evidence is based mostly on mtDNA data published during the past two decades. The major arguments in this debate have been presented at length in previous publications [9] and will not be restated herein. The most pertinent supporting material that follows will provide a foundation to the reader regarding the basics of mtDNA heredity, a review and update on the most recent mtDNA data available pertaining to the origins of Native American populations, and a summary of how this information relates to the larger DNA and Book of Mormon discussion. It is important for readers to understand that while mtDNA and other genetic motifs are useful in elucidating some historical questions, [10] it may not be possible to achieve a full resolution of questions arising between secular and religious history.
Mitochondrial DNA
The hereditary features of mtDNA provide unique information that geneticists use to study the ancient history of humanity. Such studies are based on the foundational principles of population genetics. It is essential to have a working knowledge of these principles when evaluating genetic studies relating to the Book of Mormon because those who argue against its authenticity overlook some of these concepts.
MtDNA is found in mitochondria, which are the organelles within each cell responsible for life-sustaining processes such as cell energy metabolism, cell division, and programmed cell death (apoptosis). Each cell may contain thousands of mitochondria, and each mitochondrion may contain hundreds of mtDNA genomes. A significant hereditary feature of mtDNA is that it is maternally inherited, a fact that affects the extent of historical information one can learn from its analysis.
The mtDNA molecule comprises only 16,569 bases and is therefore very small when compared to the nuclear genome (i.e., the 3.2 billion bases of genetic material that make up the twenty-three pairs of chromosomes found in the cell’s nucleus). The first complete mtDNA genome was sequenced in 1981 at Cambridge University and is called the Anderson or Cambridge Reference Sequence (CRS). [11] In 1999 Richard Andrews and colleagues resequenced the original Cambridge mtDNA, which is now referred to as rCRS. [12] This sequence became the industry standard used to compare complete or partial mtDNA data produced to date. Instead of reporting long lists of genetic bases for each mtDNA sample, a typical report includes only differences (i.e., mutations) from the rCRS. This set of mutations is called a haplotype, the mtDNA genetic profile inherited from the maternal lineage of an individual. As a general rule, mutational events occur randomly, and their accumulation over time has resulted in the differentiation of the many mtDNA lineages observed in today’s world populations. Analysis of these lineages can therefore be structured hierarchically in a treelike format called a phylogeny (fig. 1). A phylogeny attempts to model the true hereditary history of mtDNA across populations.
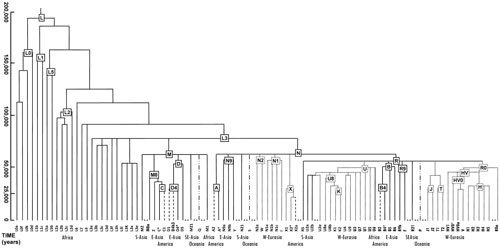
Figure 1. Schematic phylogeny of human mtDNA.
Similar to the Y chromosome (Ycs), mtDNA does not recombine with the DNA from the other molecules. That is, mtDNA is inherited as a fully intact DNA segment between generations, with variations from mother to child arising rarely due to random mutations. While the Ycs is inherited along the paternal line, as noted before, mtDNA follows an inheritance pattern found on the opposite side of the family tree, along the unbroken maternal line (fig. 2). A mother’s mtDNA is passed to all of her children, but only the daughters will pass their mtDNA to the next generation.
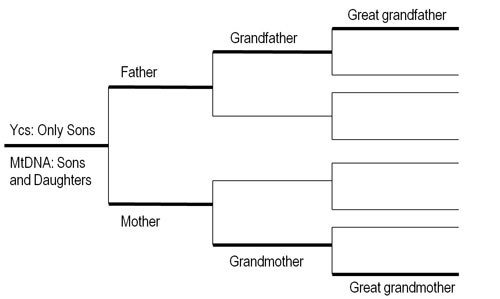
Figure 2. The paternal Y Chromosome (Ycs) and maternal mitochondrial DNA (mtDNA) inheritance patterns.
The mtDNA genome has two parts: the control region, [13] which includes three segments called HVS1, HVS2, and HVS3, [14] and the coding region (where all the mtDNA’s genes that produce proteins essential to life are found). Genetic data from an individual’s mtDNA are obtained by the following methods, with each successive approach yielding more information:
1. Inspecting restriction fragment length polymorphisms (RFLPs) using enzymes that break the DNA into smaller fragments at specific short (usually four to six base pair) sequences. Depending on the presence or lack of mutations, the fragment will or will not be broken, and the resulting fragment length indicates the presence or lack of the mutation.
2. Assaying single nucleotide polymorphisms (SNPs), where the type of base at a specific location is identified for comparison with the reference sequence.
3. Sequencing of part or the entire control region (up to approximately 1,000 bases).
4. Sequencing of the complete mtDNA genome (all the 16,569 bases—the highest level of mtDNA molecular resolution attainable).
During the 1990s, a number of studies were published presenting mtDNA data obtained from RFLP and control region sequences (often only HVS1, approximately 300 bases), many of them highlighting several Native American populations. [15] The mtDNA data produced during that decade allowed scientists to investigate for the first time the mtDNA variation from diverse populations. From this they advanced the first theories about the origin of anatomically modern humans and the processes of expansion that resulted in the colonization of the continental masses.
Starting around the year 2000, researchers employing new technological methods began to produce complete genome sequences as the standard for the most rigorous mtDNA population studies. [16] However, the process of generating a full mtDNA sequence is still labor intensive and relatively expensive. Recently, a study reviewing all the published mtDNA full sequences reported that only a very small fraction of these data are of Native American origin, leaving a considerable gap to fill in the scientific literature. [17]
The ability to acquire complete mtDNA sequences brought several benefits to the field of population genetics, including resolution of questionable phylogenies based on control region data (this region has a higher mutation rate and is therefore affected by recurring mutations); identification of smaller clades within the large world mtDNA tree; a better understanding of events that characterize the expansion and migration routes followed by our early ancestors; and an improved understanding of the expected mutation rate of the mtDNA genome, yielding a better calibration of the molecular clock—the mathematical underpinnings of historical date estimations based on genetic data.
It is important to remember that population geneticists face the continuing challenge of correlating their findings with those of other disciplines, including linguistics, anthropology, and archaeology. A multidisciplinary approach allows a consensus to be formed for date estimates and helps to cross-verify findings among different fields of study. [18]
MtDNA Haplogroups
The differentiation of mtDNA has been generated by the sequential accumulation of new mutations along radiating maternal lineages. Over the course of time, this process of molecular divergence has given rise to separate mtDNA lineages that are now called haplogroups—that is, groups of haplotypes sharing similar characteristics. Haplogroups are named following a simple but standardized nomenclature procedure, alternating letters with numbers and starting with a capital letter (e.g., K1a4, H1a, A2d2, C1b2a) (fig. 1). Coincidentally, the first time haplogroup names were given was when the sequence variation of mtDNAs from Native American populations was investigated. Four major mutational motifs were identified, and they were therefore originally named A, B, C, and D. [19]
The process of mtDNA molecular differentiation was relatively rapid and occurred mainly during and after the recent process of human colonization and diffusion into different regions and continents. Thus, serendipitously, the different subsets of mtDNA variation tend to be restricted to different geographic areas and population groups. Older mtDNA lineages had more time to accumulate a greater number of mutations, while younger mtDNA lineages accumulated fewer mutations and therefore underwent less variation. Mainstream population geneticists are in agreement that, based on the available mtDNA data, the most recent common female ancestor (from whom all mtDNAs in modern humans derive) lived in Africa about 200,000 years ago and that an initial migration out of Africa took place around 70,000 years ago, represented by an mtDNA lineage known as L3. This lineage left the Horn of Africa by migrating eastward and following a southern coastal route along the Indian Ocean, and while moving farther east about 63,000 years ago, it gave rise to two mtDNA “daughter” branches known as haplogroups M and N. An offshoot of N shortly thereafter was haplogroup R. Lineages M, N, and R are the female ancestors of all the known non-African lineages that eventually colonized the rest of the continents. These lineages are also known as macrohaplogroups or superhaplogroups. The Americas were the last of all the continents to be colonized by Homo sapiens, approximately 10,000–20,000 years ago (fig. 3).
The Basics of Population Genetics
Using the mtDNA mutations as a guide, it is possible to trace all modern mtDNA lineages back to a single African female ancestor. Geneticists have named this ancestor the African “Eve,” but despite this name, she was not necessarily the first woman on the planet. The mtDNA lineages corresponding to other women simply disappeared because their offspring failed to produce additional continuous female lineages (a phenomenon known in population genetics as genetic drift), because of natural or manmade calamities that wiped out a significant portion of the population (an event referred to as a population bottleneck), or because they were selected against due to the detrimental effect of specific mutations. This African “Eve” was the only one that was successful in perpetuating her mtDNA lineage through the generations. Therefore, because of genetic drift, population bottlenecks, and natural selection, the mtDNA lineages observed in today’s population do not reflect the full range of mtDNA variation that occurred throughout human history. A recent example from a study in Iceland based on genetic and genealogical data clearly demonstrated how the majority of people living in that country today are descended from just a small representation of people that lived there just three hundred years ago. [20] This work is a powerful illustration and a rare example of a controlled study where genealogical, historical, and genetic data are available to unequivocally demonstrate the effect of genetic drift and natural selection in a fairly isolated population. The effect of these population genetics processes occurs globally (including in organisms other than humans) and is not exclusive to the Icelandic population. Most relevant to our current discussion, these principles have also affected populations in the Americas. Although some would like to dismiss the Icelandic model and suggest that it is more an exception than the rule, [21] these population genetics laws cannot be ignored: they are the fundamental force that shaped the modern genetic landscape worldwide. It is a well-known fact that mtDNA lineages have disappeared in the past and that they will continue to disappear in modern times. This process has occurred everywhere in the world, and the Americas are no exception. [22]
Native American DNA
With regard to measuring the genetic variation observed among the indigenous people of the Western Hemisphere, molecular anthropologist Michael H. Crawford has stated this problem succinctly and repeatedly in his book The Origins of Native Americans:
The Conquest and its sequelae squeezed the entire Amerindian population through a genetic bottleneck. The reduction of Amerindian gene pools from 1/
3 to 1/ 25 of their previous size implies a considerable loss of genetic variability. . . . It is highly unlikely that survivorship was genetically random. . . . Thus, the present gene-frequency distributions of Amerindian populations may be distorted by a combination of effects stemming from genetic bottlenecks and natural selection. . . .
This population reduction has forever altered the genetics of the surviving groups, thus complicating any attempts at reconstructing the pre-Columbian genetic structure of most New World groups. [23]
Subsequent research has supported this notion. In an article dealing with ancient DNA from Native American populations that was published in the American Journal of Physical Anthropology, the authors made the following statement: “Genetic drift has also been a significant force [on Native American genetics], and together with a major population crash after the European contact, has altered haplogroup frequencies and caused the loss of many haplotypes.” [24]
These statements from experts in the field of modern and ancient DNA from Native American populations (experts not involved with the Book of Mormon and DNA debate) give insight into the influence of the major population-altering events of the Columbian and pre-Columbian eras on the genetic variation of modern Native Americans. Their mtDNA was not immune to the evolutionary processes of genetic drift and population bottleneck that have been observed in a similar fashion in other populations. One cannot overstate the importance of considering both random as well as environmental factors when studying history using DNA samples from modern populations, including that of Amerindians. Population genetics principles guide geneticists who study human history, and genetic drift and population bottlenecks are among the most basic factors considered in their work.
Some wonder if ancient DNA samples might shed additional light on the history of ancient populations such as the ancestral Native Americans. This approach can be valuable when the necessary samples are available and the DNA is of good quality. Note, however, that several limitations must be carefully considered when studying ancient DNA:
· Accessibility to the ancient remains. In many cases Native American and First Nation groups consider their burial grounds sacred and are quite resistant to DNA testing being performed on their ancestors’ remains. (Moreover, they are often resistant to testing being done on themselves.) [25]
· Contamination. Skeletal remains in museums or personal collections may have been handled improperly over time. Thus any attempt to retrieve endogenous DNA from them may be compromised by the presence of DNA belonging to those who have touched the samples since the time of their excavation.
· Confidence that the data obtained are genuine. A general practice when analyzing ancient DNA samples is to compare the data obtained with samples from the modern population. If identical or similar haplotypes are found in the modern population, then it is assumed that the data obtained from the ancient specimen are reliable. However, if no matches are found in the modern population, it can become difficult to ascertain if the data obtained belong to a lineage no longer in existence or if the genetic signal comes from contamination or postmortem damage.
· Failed sequencing due to environmental factors. Even in cases when bone fragments are found and proper excavation techniques are in place, the success rate of extracting and analyzing ancient DNA is approximately 1 in 3. Extreme heat, high humidity, gamma rays from the sun, and other factors can accelerate DNA degradation. During the last decade, thanks to new technological advancements and a better understanding of how to work with ancient DNA, [26] results have improved and the data are more reliable. However, much of the data published in the 1990s was susceptible to less rigorous collection and lab procedures that may have resulted in unreliable DNA data and conclusions.
· Limited quantity of data obtained. Because ancient DNA is highly degraded, only small fragments of genetic material can be sequenced. Most of the ancient DNA data available in the public literature comes from sequencing short segments of the control region. To date, only a few complete mtDNA sequences (the full 16,569 bases of the mtDNA genome) from ancient human remains have been successfully produced (e.g., five Neanderthals and the Tyrolean Ice Man, Ötzi). [27]
In summary, even though ancient DNA data have the potential to be extremely helpful in phylogenetic studies and in reconstructing past population events, scientists are still limited by the amount and quality of data they can obtain from ancient remains.
A significant finding that elucidates the usefulness of combining ancient and modern DNA in the study of Native American populations comes from a recent publication featuring a short control region segment sequenced from a skeleton found in Alaska that is approximately 10,000 years old. [28] Carbon dating confirmed that the remains were clearly pre-Columbian, but the genetic profile obtained did not match any of the earlier identified Amerindian mtDNAs (A2, B2, C1, D1, and X2a). Previously, a number of studies on Native American populations revealed a small quantity of samples labeled “others” since they did not belong to any of the known indigenous mtDNA lineages and were thought to have been contaminated or the result of European admixture. Based on the mtDNA data retrieved from the ancient Alaskan specimen, some of those previously unclassified samples were reexamined and are confirmed as belonging to a novel Native American lineage named D4h3. [29] Unfortunately, as explained earlier, it is difficult to access and obtain data of good quality from ancient DNA. Therefore, for every reclassified mtDNA lineage, it is probable that many misclassifications remain unknown or unresolved. The case of D4h3 is likely to be a rare event in shedding additional light on the maternal history of Native American populations.
Another serious limitation is the possibility of making inappropriate assumptions about which mtDNA candidate haplogroups to expect from the small groups described in the Book of Mormon. A survey of modern populations including Middle Easterners and Asians would reveal a certain number of mtDNA lineages that occur at high frequencies and are therefore labeled as region-specific for the modern population, but such a survey would also uncover a number of mtDNA haplogroups that are more rare. Most likely, these less frequent mtDNA lineages are the result of relatively recent migratory events, an occurrence very common throughout history because of international trade routes (such as those that took place along the Silk Road) or military expansions (e.g., the Assyrian, Babylonian, Roman, or Mongol empires). These important historical events are responsible for a partial reshuffling of the DNA compositions of geographic regions throughout the world, adding to the genetic diversity of affected locations. Although the majority of lineages found in the population of a given region display the typical mtDNA expected in that region, the reality is that potentially any given mtDNA lineage could also be found at low frequencies in the same geographic area. Any of these low-frequency haplogroups could be candidates for genetic types that were more common during an earlier time period within the last few thousand years. This issue touches on the people of The Book of Mormon because we don’t know their mtDNA affiliation. Lehi’s group could have included typical Middle Eastern lineages or rare ones, even some with a close Asian affinity. [30]
To elucidate this point, I use my own Y chromosome (Ycs) haplogroup as an example. As explained earlier, Ycs is a uniparental marker that, like mtDNA, can be traced along one specific family tree branch (in this case the direct paternal line), and for the most part it does not recombine with the other chromosomes (fig. 2). Ycs haplotypes can also be grouped in a large phylogenetic tree based on common characteristics that in most cases can be associated with specific geographic regions. I was born and raised in Italy and can trace my paternal ancestry back several generations, to the mid-seventeenth century AD. However, my Ycs belongs to haplogroup C, which has a frequency in southern Europe of less than 1 percent. Haplogroup C is found mostly in east Asia, with a branch (C4) found among the aborigines of Australia. How did haplogroup C become part of my paternal ancestry? One possibility is that it is a remnant of an ancient military expansion from the East (e.g., Mongols or Huns) that reached northern Italy. With my three sons, we contribute four instances of this particular Ycs haplotype to the population of Utah, where we currently reside. If someone took a survey of Italians in Utah with the purpose of reconstructing the typical Italian genetic composition, they would include the four of us as part of that count. This would produce a higher-than-normal frequency of haplogroup C among Utah Italians that would in turn provide a different scenario from the one observed in Italy. What if I were the first and only Italian that migrated to Utah? What is considered a rare lineage in the source population (Italy) would become the totality of the Ycs lineages for the same population in Utah. And by looking at these hypothetical data, one would likely reach the incorrect conclusion that Italians are paternally related to eastern Asian populations. This is a direct result of another principle of population genetics: the founder effect.
The same founder effect process can be observed with mtDNA lineages that are traditionally associated with Paleo-Indians who arrived in the Americas most likely via Beringia between 12,000 and 20,000 years ago. Haplogroups A2, B2, C1, and D1 are the most common mtDNAs found in Native Americans (approximately 95 percent of the population), but they do not reflect the current mtDNA landscape observed in northeast Asia. For one thing, there are no A2, B2, C1, or D1 lineages in that part of the world (one of the few exceptions is subclade C1a, found only in Asia and not in the Americas). [31] These four branches of the mtDNA world tree are exclusively found in the Americas and have been separated from all other lineages long enough to develop their own specific mutational motifs. Secondly, a survey of north Asian mtDNA lineages would reveal a much more diverse distribution and variety of mtDNA haplogroups—not a 95 percent frequency of Asian lineages belonging to subbranches of the roots A, B, C, and D. What happened to the other Asian lineages? Why are they not found in the Americas too? Genetic drift and the founder effect are again the answer. What we observe today in the Western Hemisphere are the surviving lineages that Paleo-Indians brought with them to the Americas at the time of the last ice age. The other lineages were simply lost in the process.
What about Haplogroup X?
Although the majority of mtDNA lineages surveyed to date among Amerindians belong to a subclade of one of the four Pan-American haplogroups (A2, B2, C1, and D1) having Asian affinity, this does not mean that all the pre-Columbian lineages are of Asian origins. One exception is the less common and geographically limited haplogroup X. The presence of haplogroup X in the Americas has caused no small perplexity among scientists studying Native American origins. Research questions include how haplogroup X differs from the other Pan-American haplogroups with Asian affinity, its origins, where else it is found in the world, what route it followed to the Americas, and how long ago it arrived there.
With regard to the Book of Mormon and DNA debate, haplogroup X has also played an interesting role at both ends of the spectrum in challenging or defending the historicity of the Book of Mormon. On one end are those who criticize the Book of Mormon based on the DNA data. Conversely, there are some within the LDS faith claiming that the presence of haplogroup X in the Americas supports the truthfulness of the Book of Mormon. The mutually exclusive reasoning of these two factions can be summarized as follows:
Against Book of Mormon historicity: Like other Pan-American clades, haplogroup X is of Asian origin, arriving in the Americas via Beringia (the landmass that connected northeast Siberia with modern-day Alaska during the last ice age). This migration took place more than 10,000 years ago, long before Israel ever existed.
In favor of Book of Mormon historicity: Haplogroup X is of Near Eastern origin, and its presence in the Americas represents the surviving legacy of Lehi’s party arriving in the Western Hemisphere some 2,600 years ago.
There are probably as many gradients between these two views as there are people trying to address this specific topic. However, these two points summarize most of the issues surrounding haplogroup X and the proposed association with the historicity of the Book of Mormon.
Following the discovery of the first, more common Native American mtDNA haplogroups in the early 1990s (originally termed A, B, C, and D and later renamed A2, B2, C1, and D1 to distinguish them from their Asian “cousins”), a fifth haplogroup was identified in 1996 by Peter Forster and his colleagues and named haplogroup X (not to be confused with the X chromosome). [32] Contrary to nearly all the world haplogroups, it is not geographically confined but is found at low frequency among several populations: Europeans, Africans, Asians, Middle Easterners, and Native Americans. A number of studies following the initial identification of haplogroup X among Amerindians confirmed its presence in the Western Hemisphere, [33] its variance from the X lineages found in Eurasia and Africa, and its geographic distribution confined to northern North America. [34] The Native American clade of haplogroup X is known as X2a to differentiate it from the forms of haplogroup X found in northern Africa and Eurasia. The root of this lineage is characterized by the diagnostic control region transition C16278T, and the specific X2a subclade also includes mutations at A200G and G16213A. [35]
As already discussed, the Pan-American haplogroups A2, B2, C1, and D1are clearly nested within a tree of east Asian haplogroups, thus suggesting an Asian origin followed by a Beringian migration and the differentiation of Paleo-Indian lineages from the ancestral Asian ones. However, the original differentiation of A, B, C, and D from their ancestral mtDNA lineages occurred in ancient south Asia during the early expansion of anatomically modern humans tens of thousands of years ago (south Asia is a geographic region that is not any closer to Beringia than is the Middle East). Lineages found today in central and northeast Asia (e.g., A5, B4a, C4, and D4e, to name a few) are considered “cousins” but are not ancestral to the American A2, B2, C1, and D1 haplogroups (fig. 1).
For years, scientists struggled to identify a possible Asian source for haplogroup X that could explain its presence in the Western Hemisphere. Different theories were postulated, including a possible northern Atlantic migration along the ice cap that connected northern Europe to northern America during the last ice age. This unpopular theory—referred to as the Solutrean hypothesis—was supported by archaeological discoveries revealing the presence of a similar technology in both continents arising at about the same time period. [36]
Early studies were limited to the sequence of a few hundred bases from the control region and therefore were not able to provide the level of resolution necessary to assess the phylogenetic relationship between American and Eurasian X lineages. This is particularly relevant in light of the fact that because haplogroup X initially could not be found in Asia, there was even more uncertainty regarding its origin and migration route to the Western Hemisphere. Did haplogroup X come from Europe via the glaciated northern Atlantic, or did it follow the same Beringian route as the other Native American haplogroups? If the latter was the case, why was it not found in northern Siberia or eastern Asia?
Scientists began looking for the presence of haplogroup X in other areas of Asia and eventually were able to find it in a small percentage of the Altai population, on the northern border of Mongolia. In 2001 Miroslava Derenko and his colleagues published a paper in which they suggested that intermediary positions of Altaian haplogroup X haplotypes (control region only) together with Eurasian and American X lineages and could possibly represent the population source for haplogroup X in northern North America. [37] Its absence in north Siberian populations could be explained by a rapid expansion or by genetic drift. However, when the same data were analyzed at a higher level of resolution—complete mtDNA sequences—and compared to other X haplotypes, it became evident that the Altaian mtDNA cluster (called X2e) was considerably younger than the Native American X2a. Therefore, the Asian branch of X was not ancestral to the Amerindian X2a, but it certainly could be a sister clade derived from a common, now disappeared Asian ancestor. The authors suggested that the Altaian Xs were the result of a secondary, more recent migratory event, possibly from the Caucasus region, [38] leaving the question about the origin of Native American X2a unanswered. The authors concluded their research by stating that “phylogeography of the subclades of haplogroup X suggests that the Near East is the likely geographical source for the spread of subhaplogroup X2.” [39] Interestingly, they identified a sample from Iran that shared a single, fairly conserved coding region mutation with the Native American X2a cluster: “We surveyed our Old World haplogroup X mtDNAs for the five diagnostic X2a mutations [A200G and G16213A in the control region and A8913G, A12397G, and T14502C in the coding region] and found a match only for the transition at np 12397 [nucleotide position A12397G] in a single X2* sequence from Iran. In a parsimony tree, this Iranian mtDNA would share a common ancestor with the Native American clade.” [40] However, the authors suggested that this could have been a case of IBS (identical by state, where shared mutations in different populations arise by chance in a parallel manner with no common ancestor) rather than IBD (identical by descent, where shared mutations that exist in different populations originated from a common ancestor). In other words, since they could not explain how the Iranian sample could possibly cluster with the Native American X2a lineages, they deduced that the common mutation was simply due to chance and not because of a more recent common ancestry. It wasn’t until 2008, with the publication of two papers on Middle Eastern populations, that more light was shed on the origin of haplogroup X. [41] One of them focused on the Druze population of northern Israel.
The Druze are a religious group that originated as an offshoot of Islam and now number approximately one million people living principally in Syria, Lebanon, Israel, and Jordan. The authors of the paper on Druze mtDNAs observed that most of the X lineages found elsewhere (Africa, Europe, and Asia) were also detected among the Druze, thus suggesting that they could indeed have been the source population for the spreading of haplogroup X throughout the world. Although no Native American X2a mtDNAs were observed among these people, the Altaian X2e was one of the haplotypes that the researchers identified, thus confirming a more recent migratory event that led to the presence of X2e in modern-day southern Siberia. Additionally, in 2009 a paper describing mtDNA lineages from Egyptian nomads revealed a small number of haplotypes carrying the same diagnostic coding region mutation shared by the Native American X2a samples and the one from Iran reported in 2003. [42] This finding supports the conclusion that such a mutation may indeed be ancestral to all of these samples, leaving the door open to future studies that may contribute additional knowledge about a possibly more recent (when compared to the Pan-American and Asian haplogroups) relationship between Amerindian X2a and Middle Eastern haplotypes.
This brief summary of studies focusing on the origin and diffusion of haplogroup X contains some of the details that have been used in the Book of Mormon debate over the past few years. Some Latter-day Saint scholars welcomed the association between a small group of Native American lineages and people of the Middle East as genetic evidence that indeed there was a group of seafaring Israelites that arrived in the Americas within the last couple thousand years. On the other hand, critics of the Book of Mormon dismissed this possibility by first referring to the presence of haplogroup X among the Altaians (and therefore supporting the scenario that this lineage followed the same Beringian route to the New World at the same time as the other Pan-American mtDNAs). [43] As already discussed, this first criticism is now challenged by data from complete mtDNA sequences that exclude the Asian X lineage from being the potential ancestor to the American one. A second criticism of a possible association between Book of Mormon people and the X2a lineage is based on the current coalescent age of haplogroup X2a, as well as findings based on ancient DNA studies that support a longer presence of this lineage in the Americas—close in time to the origin of other Native American haplogroups—that predates the events recorded in the Book of Mormon.
The first issue deals with the age estimate based on modern DNA. There are currently five molecular clocks that have been proposed to calculate the age of mtDNA lineages using data from coding regions or complete sequences. [44] Only one of these mutation rates is based on the complete mtDNA genome (both control and coding regions), providing an age estimate for X2a (12,800 ± 6,600 years ago), which is similar to the four Pan-American haplogroups. [45] The ages obtained using the other molecular clocks are fairly comparable. The X2a distribution limited to northern North America strongly suggests a separate migratory event from Beringia through the ice-free corridor that was open between the Cordilleran and Laurentide glaciers at the end of the last ice age. [46] In order for X2a to fit within Book of Mormon chronology, the currently accepted molecular clocks would need considerable recalibration, [47] or other samples from the Old World carrying additional mutations shared with the Native American X2a would be needed. Neither of these two scenarios is currently likely, and neither may ever become a means for conclusively demonstrating a link between X2a and Lehi’s party.
The discussion of the X haplotype illustrates the challenges encountered when attempting to reconstruct genetic scenarios from modern populations that are compatible with the Book of Mormon time line and expected source population. Based on the molecular clocks currently used by the scientific community, it would be nearly impossible to distinguish a Eurasian lineage that arrived 2,600 years ago from those brought by Europeans after the discovery of America’s double continent, simply because there would not have been enough time for these lineages to differentiate enough to allow discernment between pre-Columbian and post-Columbian admixture. The only truly testable hypothesis that unequivocally evaluates the historicity of the Book of Mormon from a molecular perspective would be to know the actual genetic profiles of Lehi’s group, identify them in the modern Native American populations, and find exact matches in samples from their Middle Eastern area of origin (assuming that genetic drift and population bottlenecks had not obliterated the genetic signal over time). Unfortunately, as already discussed, to attribute a particular genetic profile to Lehi’s group would be pure speculation, making the testing of this hypothesis impossible.
Three studies explore the presence of X2a in ancient times in the Americas. [48] As previously discussed, X2a is defined by five diagnostic mutations (two control and three coding region transitions). However, researchers of ancient mtDNA have been limited to a small segment of the control region, and therefore their classification of mtDNA lineages from ancient samples was based solely on one basal mutation for the root of haplogroup X (C16278T). This mutation is shared by all the X lineages worldwide and is also a mutational hotspot—a nucleotide position that recurrently mutates in the world mtDNA phylogeny. According to a recent publication surveying 2,000 complete mtDNA sequences, C16278T was the twelfth most common mutation observed. [49] Using this single site as the diagnostic mutation to place ancient samples into the X haplogroup already poses a serious limitation to the accuracy of such inferences. Two of the three papers in question predate the era of gold standards for ancient DNA studies, which already constitutes a potential concern in confidently accepting the quality of their results and subsequently of the conclusion derived from such analyses.
The first paper dealt with a burial site in Windover, Florida, where 176 individuals were identified and recovered. The site was carbon-dated approximately seven to eight thousand years ago, and a short section of the mtDNA control region (166 bases from position 16151 to position 16317) was sequenced for fourteen individuals. Two of the specimens analyzed yielded the recurrent mutation C16278T, which is also diagnostic for the whole X haplogroup. However, neither of them reported the common G16213A transition, which would have been found within the range that was sequenced. Additionally, the mutations of these two haplotypes are not sufficient to allow an unambiguous assignment to either haplogroup X2a or any of the other Pan-American haplogroups. The authors admitted that “given the limited number of Windover samples currently analyzed and the restricted length of mtDNA sequences analyzed . . . any inference regarding Windover structure or its relationship to contemporary Amerind groups is necessarily tentative.” [50]
The second paper dealing with haplogroup X2a from ancient DNA was based on specimens retrieved from a Native American cemetery at the Norris Farm site in Illinois. Archaeologists classified the site as being part of the Oneota culture and dated it at about one thousand years ago, a time frame that would somewhat fit with Book of Mormon chronology. DNA was extracted successfully from 108 individuals, but only 52 of them were sequenced for a segment of the mtDNA control region (353 bases, from position 16056 to position 16409). Nearly all haplotypes were assigned to one of the four major Pan-American haplogroups, with the exception of two that bore the X-specific C16278T transition and none of the A2, B2, C1, and D1 diagnostic mutations. However, as with the previous paper, both samples are missing mutation G16213A, which is found in nearly all modern-day X2a samples.
Contamination, postmortem damage, parallel or back mutations, and misclassification due to the limited data available might explain the presence of C16278T and the absence of G16213A, which precluded a confident determination of haplogroup X in the ancient burial sites described in these first two papers. [51] Currently, the first convincing evidence of haplogroup X in pre-Columbian America comes from a 2002 study reporting ancient DNA data from an approximately 1,340-year-old burial site on the Columbia River near Vantage, Washington. [52] The possibility that future studies—including a more detailed and controlled analysis of the samples from the 8,000-year-old Windover burial site—may eventually confirm the presence of haplogroup X in pre–Book of Mormon America is not excluded. It is also possible that the specimens analyzed could belong to a rarer or extinct X subclade, distantly related to the more common X2a found in the modern native population of northern North America, as demonstrated by the recently discovered X2g lineage found in an Ojibwa sample. [53]
What about Other mtDNA Lineages Found in the Americas?
Molecular anthropologist Ted Schurr addressed the issue of Amerindian lineages not belonging to the classical Pan-American haplogroups by stating that
Various studies have also revealed a high frequency of “private haplotypes” in individual populations or groups of related Amerindian tribes. These patterns reflect the role that genetic drift and founder effects have played in the stochastic extinction and fixation of mtDNA haplotypes in Native American populations.
A number of haplotypes not clearly belonging to these five maternal lineages have been also detected in different Native American groups. These “other” mtDNAs have often been considered additional founding haplotypes or haplogroups in New World populations. However, most have since been shown to be derivatives of haplogroups A–D that have lost diagnostic mutations. The remainder appears to have been contributed to indigenous groups through nonnative admixture. In addition, the “other” mtDNAs detected in archeological samples may have resulted from contamination with modern mtDNAs, or were insufficiently analyzed to make a determination of their haplogroup status. [54]
The process of discovering additional pre-Columbian lineages in the Americas is somewhat limited by the preconceived notion that if a lineage does not fit with the classical Native American haplotypes, it is most likely the result of a recent migratory event from the Old World. For example, a 1999 study on the Cayapa tribe of Ecuador revealed a number of lineages that did not fit with the five known Native American haplogroups. [55] Although the authors believed it could have been a newly identified pre-Columbian lineage and called it the “Cayapa haplotype,” others dismissed it as a possible case of European mtDNA introgression. [56] However, it was only when mtDNA data became available from the approximately 10,000-year-old Alaskan skeleton described earlier that the Cayapa haplotype was confirmed as a genuine pre-Colombian novel lineage. [57] From the initial four Amerindian mtDNA haplogroups discovered in the early 1990s, at least fifteen Native American founding lineages are currently catalogued, and it is very likely that more will be identified in future studies. [58] Additionally, detailed studies based on complete mtDNA sequences of haplotypes belonging to peculiar branches of the four Pan-American haplogroups may reveal interesting distribution patterns reflecting novel migratory events that could not be detected based on control region data only.
Intriguingly, two recent papers about mtDNA variation in Northern Asia [59] and in Icelanders [60] reinforced the difficulty in attempting a one-size-fits-all explanation of the peopling of the Americas. In the first study, the authors analyzed 770 samples, mostly from Eastern and Northern Asia and belonging to mtDNA haplogroups C and D. Although these two lineages are also found at high frequencies among American natives, a dissection at the subclade level reveals a clear distinction and no overlap between Asian and American haplotypes. In fact, if we take haplogroup C as an example, subclades C1a, C4, C5, and C7 were found in the Asian dataset, none of which have been detected in the Americas, which instead is characterized only by the presence of three separate C1 branches, namely C1b, C1c, and C1d. [61] An additional C lineage (C4c) has also been confirmed as an autochthonous Amerindian haplotype. [62] While all these lineages are considered sister clades, the geographical origin and dispersal of their ancient female ancestor (haplogroup C) is largely unknown. A similar outcome was observed in the study of haplogroup D, with the significant finding of a potentially novel northern North American lineage called D4b1a2a1a. The lack of overlap between Asian and American C and D haplogroups could be explained by the random disappearance of certain haplotypes due to genetic drift, as it was explained earlier. However, if this is the case, the same factors could certainly have affected the survival, distribution, and frequency of any other lineage that arrived in the Americas in ancient or recent times.
The second study probably brings more questions than answers to the table. The authors reported a fifth C1 lineage found in Iceland. How does it fit with the rest of the mtDNA C1 picture? Genealogical and phylogenetic analyses of the Icelandic C1 haplotype (called C1e) revealed that its presence in Iceland is indeed quite ancient, and not the result of recent admixture, as it was initially postulated. The Icelandic C1e is characterized by a set of 11 control and coding region mutations that distinguish it from its four sister clades. A survey of worldwide mtDNA samples (both complete sequences and control region haplotypes in modern and ancient mtDNAs) identified a single sample sharing C1e characteristics in a German individual. Because of its rarity in Western Eurasia, the authors excluded the Old World as the cradle for the novel Icelandic mtDNA. However, there is enough evidence to conclude that such lineage was present in the Icelandic mtDNA pool prior to Christopher Columbus’ arrival in the Americas, thus raising the possibility of a pre-1492 contact between Icelanders and American natives. The already-mentioned Solutrean Hypothesis (which is highly disputed and controversial) [63] could be resurrected due to the discovery of this new lineage. However, the authors feel more comfortable with the possibility of Viking voyages between Iceland and North America which resulted in the migration of at least one Native American woman (carrying the C1 lineage) back to the Old World. Since no C1e lineage has been detected in modern American indigenous populations, the authors emphasized that “given the rather drastic population size reductions that resulted from the actions of Europeans after 1492, it is quite possible that the C1e lineage was once carried by, but has now been lost from, contemporary individuals with Native American matrilineal ancestry.” [64]
This discovery has a number of implications for the ongoing debate regarding DNA and the Book of Mormon:
1. We are reminded once more that we are dealing with related sister clades, but we still don't know much about their ancestral nodes. In other words, we know that C1a is found exclusively in Asia, that C1b, C1c, and C1d are indigenous mtDNA lineages of the Americas, and now we know that C1e is found in the Old World and nowhere else. However, no traces of the ancestral C1 haplogroup in modern populations has been found, thus leaving us with an estimate age for its existence (approximately 20,000 years ago), [65] but not an exact geographic location for its origin. If the founders of two populations originally co-existed in the same region and then both migrated somewhere else and founded a population in their new regions, we cannot necessarily assume that they lived in the same place – all we can know is only how far back in time that common ancestor lived. If Beringia was once the refuge for the ancestors of all the C1 mtDNA lineages known today, then the only explanation is that C1e eventually found its way to Iceland leaving no trace of its presence in the American continent. This is a plausible hypothesis, but it definitely leaves room for additional explanations.
2. If C1e is instead an Old World mtDNA lineage (a sister clade distantly related to the Asian-American ones), then a similar argument could be proposed for haplogroup X2a, which is also found exclusively in the Americas and it is quite different from its Old World counterparts (e.g. X1, X2b, X2c, X2d, X2e, etc.). More specifically, the same logic used in promoting X2a as a marker with a strong Near Eastern affinity should also apply for the newly discovered C1e lineage, whose related lineages are found all over North, Central, and South America. There is no doubt that all these lineages share a common ancestor and are related, but we are unsure about when and especially where their female progenitors lived and how these clades expanded from their original geographic areas to their current locations.
To summarize, it appears that as new mtDNA lineages with Native American affinity are identified and made available through scholarly publications, the question surrounding the peopling of the Americas and the contacts these Natives may have had through the millennia with Old World migrants is far from completely resolved. As the authors of the Icelandic study wrote, “…although the complete sequence of the Icelandic C1 lineage fully resolved its position in the human mtDNA tree, the mystery surrounding the lineage's geographical origin has only deepened.” [66] This is basically the case for all Native American mtDNA clades. The risk of the critics' premature dogmatism is evident.
What about Other Genetic Markers?
The purpose of this essay is to provide an updated review of mtDNA research on Native American populations in light of the Book of Mormon debate. In the interest of space, it is not feasible to discuss in detail data from additional genetic systems, but a brief review of findings will be highlighted.
The Ycs data produced to date are still fairly scarce and have produced discrepant results, suggesting that considerable work to fully investigate the history of paternal lineages in the Americas is still badly needed. Future studies will need to test many more samples at a higher level of resolution in order to achieve a greater dissection and understanding of Amerindian Ycs haplogroups, including a better calibration of the Ycs molecular clock. Additionally, while autochthonous mtDNA lineages are still found abundantly among both indigenous and mixed American populations, [67] the European male contribution to the Native American gene pool was devastating in terms of preserving the Native American genetic signal. The genetic bottleneck experienced in Ycs lineages was tenfold more severe than in its female counterpart, thus making studies based on this uniparental paternal marker far less informative in elucidating Native American genetic history.
With regard to autosomal DNA, the genetic reshuffling that occurs in each generation creates serious limitations in the ability to trace specific ancestral lineages unequivocally. When compared to Ycs and mtDNA markers, the study of autosomal DNA is far more complex and is less forthcoming in straightforward interpretation. Recent technological advances now allow for the testing of up to one million polymorphic autosomal sites for an individual, providing an unprecedented level of resolution in characterizing an individual’s genetic profile. From such an abundance of data, statistical analysis can give the estimated percentage of an ancestral population’s contribution to an individual’s genetic makeup. This can provide a picture of possible genetic influences from other populations that may not be reflected in the strictly maternal or paternal ancestral lineage. However, with regard to the Book of Mormon discussion, autosomal DNA inheritance is subject to the same population forces as other genetic systems (genetic drift, genetic bottleneck, and founder effect), and considering the likely demographic scenario of the Book of Mormon (i.e., a small group of Old World migrants mixing with a large population of ancient Asian origins), the probable findings of autosomal studies are unlikely to contradict results already achieved with mtDNA and Ycs data. Native American Ycs, mtDNA, or autosomal DNA data analyzed will likely continue to produce a predominantly Asian signal.
A recent study based on a small section of DNA found on chromosome 9 attempted to determine the origin and number of Paleo-Indian migrations. Based on their analysis, the authors concluded that “all modern Native Americans . . . trace a large portion of their ancestry to a single founding population that may have been isolated from other Asian populations prior to expanding into the Americas.” [68] This study was recently mentioned as further demonstration that conclusions by critics of the Book of Mormon in the past are indeed correct, based on the fact that the study purportedly reported that all Native American populations and all individuals analyzed carried the same autosomal polymorphic mutation also found in Asian populations. [69] Book of Mormon opponents, however, disregard several key points in their arguments. First, as already discussed, the presence of indigenous populations of Asian origins prior to the arrival of Book of Mormon people does not affect the historicity of the book itself. These autosomal findings are in line with what is already known about Native American populations and do not change arguments already presented that propose that Book of Mormon events are compatible with the Asian-dominated genetic landscape found in Native Americans today. Population bottleneck, founder effect, genetic drift, and other population-altering forces affect all genetic systems, including autosomal DNA. It would not be unusual to expect that the small autosomal contribution of Lehi and his followers could be lost over time when mixing with an already established population of Asian origin. Additionally, the authors concluded that “a large portion” and not all the Native American ancestry can be traced to a single population with Asian affinity. A further important point comes from the idea of hypothesis construction. This research was not designed to identify a possible presence of Western Eurasian–specific markers in the Amerindian populations, and thus it is not surprising that none were found.
Of greater relevance to the debate about possible subsequent migrations to the Western Hemisphere, besides those that occurred after the last ice age, is a recent study published in the prestigious journal Nature. The authors reported autosomal DNA data that were successfully sequenced from hair belonging to a well-preserved 4,000-year-old Saqqaq individual discovered in Greenland. [70] This research has contributed greatly to the current understanding of events that led to the peopling of the Americas. The authors concluded that the genetic makeup of the ancient Saqqaq individual was very different from that of Inuit or other Native American populations. Instead, he was closely related to Old World Arctic populations of the Siberian Far East, being separated from them by approximately two hundred generations (roughly 5,500 years). These data suggest a distinctive and more recent migration across Beringia by a group of people that were not related to the ancestors of modern-day Native Americans, who arrived on the American continent nearly 10,000 years earlier. As the senior author emphasized, the lack of genetic continuity between the ancient Saqqaq individual and the modern population of the New World Arctic stands as a witness that other migrations that left no contemporary descendants could have taken place. [71] In commenting about the findings of this project, population geneticist Marcus Feldman from Stanford University said that “the models that suggest a single one-time migration are generally regarded as idealized systems, like an idealized gas in physics. But there may have been small amounts of migrations going on for millennia.” He went to explain that “just because researchers put a date on when ancient humans crossed the Bering Bridge, that doesn’t mean it happened only once and then stopped.” [72] Moreover, a multiple population source/
Lastly, an important contribution to the field of anthropological genetics has been the publication in 2010 of the first-draft sequence of a Neanderthal genome. [74] This truly remarkable achievement was also noted in an entry under the title of “Neanderthals and Lamanites: New Science Impacts Book of Mormon DNA Studies” found on the Signature Books website, [75] in which the author praised the new technological advancement in the field of autosomal DNA testing, which now allows the sequencing of large amounts of SNP data obtained from both modern and ancient specimens. Consistent with the data reported, the Signature Books author emphasized that now “we have learned that humans and Neanderthals bred with each other 30,000 years ago.” However, what he/
We detect gene flow from Neandertals into modern humans but no reciprocal gene flow from modern humans into Neandertals. Although gene flow between different populations need not be bidirectional, it has been shown that when a colonizing population (such as anatomically modern humans) encounters a resident population (such as Neandertals), even a small number of breeding events along the wave front of expansion into new territory can result in substantial introduction of genes into the colonizing population (emphasis added).
As the authors point out, natural selection plays a determinant role in the successful propagation of genes to future generations. In this case, Homo sapiens was successful in physically removing all the competitors, or else the stronger genetic adaptation and fitness of the resident population would have eventually won the “genetic battle” against the new comers, as witnessed by the small percentage of Neanderthal DNA that surivived in modern humans. This effect is very common among migrating human populations, as it is for other species. It is highly unlikely that a small group of people arriving to an already-occupied territory and inbreeding with the local population, would be successful in perpetuating a genetic legacy that would have survived to modern times. Just as DNA from Neanderthals is found in anatomically modern humans (and not vice versa), it is no surprise that the DNA of all the modern-day Native Americans carries the genetic signal of their ancestral resident population (which shared an ancient Asian origin), instead of pre-Columbian Old World DNA brought by Lehi and his group.
Conclusions
The Book of Mormon is not a volume about the history and origins of all American Indians. A careful reading of the text clearly indicates that the people described in the Book of Mormon were limited in the recording of their history to events that had religious relevance and that occurred in relatively close proximity to the keepers of the annals.
The fact that the DNA of Lehi and his party has not been detected in modern Native American populations does not demonstrate that this group of people never existed or that the Book of Mormon cannot be historical in nature. The absence of evidence is not evidence of absence. [76] Further, the very idea of locating the genetic signature of Lehi’s family in modern populations constitutes a truly untestable hypothesis since it is not possible to know the nature of their genetic profiles. Without our knowing the genetic signature to be located, any attempt at researching it will unavoidably result in further assumptions and untestable hypotheses. What were the characteristics of Lehi’s DNA and the DNA of those who went along with him? What haplogroup(s) did they belong to? We will never know. Yet this key point seems lost on those who insist on using genetic evidence as a means to validate or reject the Book of Mormon as a historical narrative. Attempting to make such conclusions is a miscarriage of logic comparable to collecting and analyzing the DNA of thousands of people living in the area surrounding a hypothetical crime scene from which no DNA could be retrieved from the individual who committed the crime, thus creating a comprehensive database of all these people. Will the database include the DNA signature of the criminal? If so, how could the perpetrator be identified among the thousands of others? Similarly, would a database composed of thousands of Native American DNA samples provide the necessary evidence to validate the existence of a small group (perhaps as few as two mtDNA haplotypes) that migrated from the Old World and settled somewhere in the Americas? Conversely, could haplogroup X be undoubtedly inferred as the ultimate proof of the genetic legacy this group left, without ever knowing their actual original DNA signature? Mitochondrial DNA is a powerful tool in reconstructing the history of our race, as demonstrated by the numerous publications that have been produced over the past two-and-a-half decades. However, as has been amply demonstrated, knowing a great deal about the genetic composition of modern-day Native American populations does not give conclusive evidence of the validity or the implausibility of the Book of Mormon’s historicity.
An additional caveat is the lack of professional training in population genetics by those promoting a supposed discrepancy between the genetic evidence and the Book of Mormon account. Some of them claim that their conclusions are strongly supported by trained experts who have been consulted for unbiased opinions about this particular matter. [77] This should raise some concerns, though, since it is fairly obvious that most people outside of the circle of Mormonism have very limited knowledge of the Book of Mormon and its contents. As a further counterpoint to the critics’ arguments, these experts seem to be in agreement that DNA lineages from a small Old World group migrating to an already heavily populated American continent would disappear. [78] Moreover, it is also noteworthy that what these scientists know about what Latter-day Saints believe has been provided mainly as one-sided background information from the critics themselves. To offer a personal anecdote, my scientist colleagues have asked me about DNA evidence and the Book of Mormon on several occasions. I respond with a simple summary in which I explain that the DNA lineages of Lehi’s colony could have been lost due to genetic drift since the number of people involved was probably fairly small compared to the size of the resident Amerindian population. I also explain that it is not possible to distinguish those lineages from post-Columbian admixture, simply because 2,600 years is not enough time for Book of Mormon mtDNA to differentiate Lehi’s descendants from their Eurasian counterparts. My colleagues typically reply that they are not convinced that I have accurately represented what Latter-day Saints believe—namely, that Lehi’s posterity comprises all Native Americans. These personal experiences give context for evaluating “genuine experts’ opinions,” based as they are on what the critics may have shared as background information regarding the Book of Mormon and Latter-day Saint beliefs. Ultimately, the critics’ arguments hold up only when they prescribe what it is that Latter-day Saints believe. Since neither the Book of Mormon nor Church doctrine indicates that all Native Americans descend from the Book of Mormon people, the critics’ arguments are on a weak footing at the outset.
In light of the information provided in this essay, it should be evident that the work of reconstructing the history of Native American populations using molecular data is still under way. Some questions can be answered, but many more remain, and spur further research. The genetic evidence of the peopling of the Americas is not fully understood, and it has evolved substantially over the past two decades. DNA research, and particularly mtDNA data, has been produced in great abundance during this time period and has provided an initial glimpse into the history and prehistory of the indigenous peoples of the Western Hemisphere. This is truly an exciting time to study the genetic history of Native Americans, for there is much yet to be understood. For example, how is the high frequency of haplogroup B in Southeast Asia and western South America reconciled with its rarity in the native populations of north Siberia and Alaska? The scarcity of archaeological evidence for human settlements on either side of the Bering Strait provides a degree of intrigue, considering that mainstream scientists currently accept Beringia as the likely refugium for Paleo-Indians during the last ice age, leaving open the possibility for alternative routes into the Americas. [79]
Mitochondrial DNA is doubtless a powerful tool that can reveal details about the expansion processes leading to the colonization of the world, including America’s double continent. However, it is not well suited as the ultimate tool to assess the historicity of religious documents like the Book of Mormon and the Bible. If the DNA of Lehi and his family cannot be confidently detected in the modern Amerindian population, does it mean that they never existed? The principles underlying this question can be further extrapolated to other religious scenarios. Can we use DNA to decisively prove that the great biblical patriarchs—Abraham, Isaac, and Jacob—ever existed? What were their own and their descendants’ mtDNA haplotypes? What about the other great Old Testament figures, such as Joseph of Egypt, Moses, and Isaiah? Can we use DNA analysis to prove that Jesus Christ lived? The New Testament mentions that Jesus had brothers and sisters (Matthew 13:55–56; Mark 6:3) through whom Mary’s mtDNA could have been transmitted to future generations (and if not through Mary, perhaps through some of her female relatives). Where is their DNA in today’s population? Would it be acceptable to conclude that these are fictional historical figures and the biblical text a hoax because of the lack of genetic evidence?
As I already commented on another occasion, “I find no difficulties in reconciling my scientific passion about Native American history with my religious beliefs. I am not looking for a personal testimony of the Book of Mormon in the double helix. The scientific method and the test of faith are two strongly connected dimensions of my existence, working synergistically in providing greater understanding, knowledge, and from time to time even a glimpse into God’s eternal mysteries.” [80] Anyone using DNA to ascertain the accuracy of historical events of a religious nature—which require instead a component of faith—will be sorely disappointed. DNA studies will continue to assist in reconstructing the history of Native American and other populations, but it is through faith that we are asked to search for truth in holy writings (Moroni 10:3–5). [81]
Notes
This chapter is an updated version of an article published in FARMS Review 22, no. 1 (2010): 191–227. I am grateful to the following individuals for commenting on this manuscript: Dr. Alessandro Achilli (University of Perugia, Italy), Jayne E. Ekins, Diahan Southard, and Dr. Scott R. Woodward (Sorenson Molecular Genealogy Foundation, USA), Professor Antonio Torroni (University of Pavia, Italy), Dr. Amy Williams (Harvard Medical School, USA), and Dr. Gregory L. Smith (University of Calgary, Canada).
[1] Dennis H. O’Rourke, “Human Migrations: The Two Roads Taken,”Current Biology19, no. 5 (2009): R204.
[2] Michael Crawford,The Origins of Native Americans: Evidence from Anthropological Genetics(Cambridge: Cambridge University Press, 1998), 2.
[3] Geraldine Barnes,Viking America: The First Millennium(Suffolk, England: St. Edmundsbury Press, 2001). Note that no genetic contribution from Vikings has been detected to date in the modern Native American population. Either they kept to themselves and were not welcomed by native groups, or their DNA has not yet been identified in contemporary Amerindians. John L. Sorenson, “Ancient Voyages Across the Ocean to America: From ‘Impossible’ to ‘Certain,’”Journal of Book of Mormon Studies14, no.1 (2005): 6, notes that the Viking presence in North America has been considered to be of no historical importance and goes on to present “decisive” empirical evidence of transoceanic distribution of flora and fauna in pre-Columbian times. See also Martin H. Raish and John L. Sorenson,Pre-Columbian Contacts with the Americas across the Oceans: An Annotated Bibliography, 2 vols. (Provo, UT: Research Press, 1996).
[4] Antonio Torroni and others, “Asian Affinities and Continental Radiation of the Four Founding Native American mtDNAs,”American Journal of Human Genetics53, no. 3 (1993): 563–90; and Alessandro Achilli and others, “The Phylogeny of the Four Pan-American MtDNA Haplogroups: Implication for Evolutionary and Disease Studies,”PloS ONE3, no. 3 (2008): e1764.
[5] For a summary of the principal theories of Book of Mormon New World geography, see http://
[6] Carrie A. Moore, “Debate renewed with change in Book of Mormon introduction,”Deseret Morning News, www.deseretnews.com/
[7] In Conference Report, April 1929, 15–16.
[8] See, for example, John L. Sorenson, “When Lehi’s Party Arrived in the Land, Did They Find Others There?”, Journal of Book of Mormon Studies1 (1992): 1–34; John L. Sorenson and Matthew Roper, “Before DNA,”Journal of Book of Mormon Studies12 (2003): 6–23; and Blake T. Ostler, “DNA Strands in the Book of Mormon,”Sunstone, May 2005, 63–71.
[9] This issue has been dealt with competently in Daniel C. Peterson, ed.,The Book of Mormon and DNA Research(Provo, UT: Neal A. Maxwell Institute for Religious Scholarship, 2008). Examples of Book of Mormon criticisms based on alleged DNA evidence are found in Simon G. Southerton,Losing a Lost Tribe: Native Americans, DNA, and the Mormon Church(Salt Lake City: Signature Books, 2004); Thomas W. Murphy, “Lamanite Genesis, Genealogy, and Genetics,” inAmerican Apocrypha: Essays on the Book of Mormon, ed. Dan Vogel and Brent L. Metcalfe (Salt Lake City: Signature Books, 2002), 47–77; and Brent L. Metcalfe, “Reinventing Lamanite Identity,”Sunstone, March 2004, 20–25. A seriously flawed attempt by a nonspecialist to adduce DNA evidence in favor of Book of Mormon historicity is Rod L. Meldrum,Rediscovering the Book of Mormon Remnant through DNA(Honeoye Falls, NY: Digital Legend Press, 2009).
[10] See, for example, Ugo A. Perego, Jayne E. Ekins, and Scott R. Woodward, “Mountain Meadows Survivor? A Mitochondrial DNA Examination,”Journal of Mormon History32, no. 3 (Fall 2006): 45–53.
[11] Stephen Anderson and others, “Sequence and Organization of the Human Mitochondrial Genome,”Nature290 (1981): 457–65.
[12] Richard M. Andrews and others, “Reanalysis and Revision of the Cambridge Reference Sequence for Human Mitochondrial DNA,”Nature Genetics23, no. 2 (1999): 147.
[13] Also called the hypervariable or D-loop region.
[14] Sometimes referred to as HVR1, HVR2, and HVR3.
[15] For example, Antonio Torroni and others, “Native American Mitochondrial DNA Analysis Indicates That the Amerind and the Nadene Populations Were Founded by Two Independent Migrations,”Genetics130 (1992): 153–62; Antonio Torroni and others, “mtDNA and Y-Chromosome Polymorphisms in Four Native American Populations from Southern Mexico,”American Journal of Human Genetics54, no. 2 (1994): 303–18; Antonio Torroni and others, “Mitochondrial DNA ‘Clock’ for the Amerinds and Its Implications for Timing Their Entry into North America,”Proceedings of the National Academy of Sciences91, no. 3 (1994): 1158–62; and Peter Forster and others, “Origin and Evolution of Native American mtDNA Variation: A Reappraisal,”American Journal of Human Genetics59 (1996): 935–45.
[16] Antonio Torroni and others, “Do the Four Clades of the mtDNA Haplogroup L2 Evolve at Different Rates?”American Journal of Human Genetics69, no. 6 (2001): 1348–56.
[17] Luísa Pereira and others, “The Diversity Present in 5140 Human Mitochondrial Genomes,”American Journal of Human Genetics84 (2009): 628–40; and Mannis van Oven and Manfred Kayser, “Updated Comprehensive Phylogenetic Tree of Global Human Mitochondrial DNA Variation,”Human Mutation30, no. 2 (2009): E386–94, www.phylotree.org. As of March 1, 2011, the publicly accessible GenBank database contains 8,731 complete mtDNA sequences, but known Native American haplogroups still suffer from significant underrepresentation. See also www.ncbi.nlm.nih.gov/
[18] Alessandro Achilli and Ugo A. Perego, “Mitochondrial DNA: A Female Perspective in Recent Human Origin and Evolution,” inOrigins as a Paradigm in the Sciencesand in the Humanities, ed. Paola Spinozzi and Alessandro Zironi (Goettingen: V&R unipress, 2010), 41–58.
[19] Torroni, “Asian Affinities.”
[20] Agnar Helgason and others, “A Populationwide Coalescent Analysis of Icelandic Matrilineal and Patrilineal Genealogies: Evidence for a Faster Evolutionary Rate of mtDNA Lineages than Y Chromosomes,”American Journal of Human Genetics72, no. 6 (2003): 1370–88.
[21] Simon Southerton, “Answers to Apologetic Claims about DNA and the Book of Mormon,” www.irr.org/
[22] http://
[23] Crawford,Origins of Native Americans, 49–51, 239–41, 260–61.
[24] Beth A. S. Shook and David G. Smith, “Using Ancient mtDNA to Reconstruct the Population History of Northeastern North America,”American Journal of Physical Anthropology137 (2008): 14.
[25] Amy Harmon, “DNA Gatherers Hit Snag: Tribes Don’t Trust Them,”New York Times, December 10, 2006, www.nytimes.com/
[26] Alan Cooper and Hendrik N. Poinar, “Ancient DNA: Do It Right, or Not at All,”Science289, no. 5482 (2000): 1139.
[27] Adrian W. Briggs and others, “Targeted Retrieval and Analysis of Five Neandertal mtDNA Genomes,”Science325 (2009): 318–21; and Luca Ermini and others, “Complete Mitochondrial Genome Sequence of the Tyrolean Iceman,”Current Biology18 (2008): 1687–93.
[28] Brian M. Kemp and others, “Genetic Analysis of Early Holocene Skeletal Remains from Alaska and Its Implications for the Settlement of the Americas,”American Journal of Physical Anthropology132 (2007): 605–21.
[29] Ugo A. Perego and others, “Distinctive Paleo-Indian Migration Routes from Beringia Marked by Two Rare MtDNA Haplogroups,”Current Biology19, no. 1 (2009): 1–8. A single haplotype sharing part of the D4h3 motif was also identified in the province of Shandong, China, out of more than 10,000 Asian mtDNAs.
[30] Although some information is available about the ancestry of Lehi and Ishmael, we know nothing about the origins of Sariah and Ishmael’s wife, who were responsible for passing their mtDNA to future generations.
[31] Erika Tamm and others, “Beringian Standstill and Spread of Native American Founders,”PloS ONE2, no. 9 (2007): e829.
[32] Forster, “Origin and Evolution of Native American mtDNA Variation.”
[33] Michael D. Brown and others, “MtDNA Haplogroup X: An Ancient Link between Europe/
[34] Rosaria Scozzari and others, “MtDNA and Y Chromosome-Specific Polymorphisms in Modern Ojibwa: Implications about the Origin of Their Gene Pool,”American Journal of Human Genetics60, no. 1 (1997): 241–44; and Perego, “Distinctive Paleo-Indian Migration.”
[35] Achilli, “The Phylogeny of the Four Pan-American MtDNA Haplogroups”; and Perego, “Distinctive Paleo-Indian Migration.”
[36] Dennis Stanford and Bruce Bradley, “Ocean Trails and Prairie Paths? Thoughts about Clovis Origins,” inThe First Americans: The Pleistocene Colonization of the New World, ed. Nina G. Jablonski (San Francisco: Academy of Science, 2002), 255–71; and Bruce Bradley and Dennis Stanford, “The North Atlantic ice-edge corridor: a possible palaeolithic route to the New World,”World Archaeology36, no. 4 (2004): 459–78.
[37] Miroslava V. Derenko and others, “The Presence of Mitochondrial Haplogroup X in Altaians from South Siberia,”American Journal of Human Genetics69, no. 1 (2001): 237–41.
[38] Maere Reidla and others, “Origin and Diffusion of mtDNA Haplogroup X,”American Journal of Human Genetics 73, no. 5 (2003): 1178–90.
[39] Ibid.
[40] Ibid.
[41] Shlush and others, “The Druze: A Population Genetic Refugium of the Near East,”PloS ONE3, no. 5 (2008): e2105; and Doron M. Behar and others, “Counting the Founders: The Matrilineal Genetic Ancestry of the Jewish Diaspora,”PloS ONE3, no. 4 (2008): e2062.
[42] Martina Kujanová and others, “Near Eastern Neolithic Genetic Input in a Small Oasis of the Egyptian Western Desert,”American Journal of Physical Anthropology140, no.2 (2009): 336–46.
[43] Perego, “Distinctive Paleo-Indian Migration.”
[44] For details about the five age-estimate models based on complete mtDNA sequences, see Dan Mishmar and others, “Natural selection shaped regional mtDNA variation in humans,”Proceedings of the National Academy of Sciences100, no. 1 (2001): 171–76; Toomas Kivisild and others, “The Role of Selection in the Evolution of Human Mitochondrial Genomes,”Genetics172, no. 1 (2006): 373–87; Perego, “Paleo-Indian Migration”; Pedro Soares and others, “Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock,”American Journal of Human Genetics84, no. 6 (2009): 740–59; and Eva-Liis Loogväli and others, “Explaining the Imperfection of the Molecular Clock of Hominid Mitochondria,”PloS ONE4, no. 12 (2009): e8260.
[45] Soares, “Correcting for Purifying Selection.”
[46] Perego, “Distinctive Paleo-Indian Migration.”
[47] Supporters of X haplogroup as evidence for Book of Mormon historicity and its geographic setting in northern North America rely on the unpopular molecular clock proposed by a forensic team in 1997. This clock was based on control region data only. See Thomas J. Parsons and others, “A high observed substitution rate in the human mitochondrial DNA control region,”Nature Genetics15 (1997): 363–68.
[48] William W. Hauswirth and others, “Inter- and Intrapopulation Studies of Ancient Humans,”Experientia50, no. 6 (1994): 585–91; Anne C. Stone and Mark Stoneking, “MtDNA Analysis of a Prehistoric Oneota Population: Implications for the Peopling of the New World,”American Journal of Human Genetics62, no. 5 (1998): 1153–70; and Ripan S. Malhi and David G. Smith, “Brief Communication: Haplogroup X Confirmed in Prehistoric North America,”American Journal of Physical Anthropology119 (2002): 84–86.
[49] Soares, “Correcting for Purifying Selection.” A similar outcome was observed when querying the Sorenson Molecular Genealogy Foundation mtDNA database (www.SMGF.org). Out of more than 76,000 samples, C16278T was observed in 8,501 cases in several haplogroups, including all the Pan-American lineages.
[50] Hauswirth, “Inter- and Intrapopulation Studies of Ancient Humans.”
[51] Malhi and Smith, “Haplogroup X Confirmed.”
[52] Malhi and Smith, “Haplogroup X Confirmed.”
[53] Perego, “Distinctive Paleo-Indian Migration.”
[54] Theodore G. Schurr, “The Peopling of the New World: Perspectives from Molecular Anthropology,”Annual Review of Anthropology33 (2004): 556.
[55] Olga Rickards and others, “MtDNA History of the Cayapa Amerinds of Ecuador: Detection of Additional Founding Lineages for the Native American Populations,”American Journal of Human Genetics65, no. 2 (1999): 519–30.
[56] Schurr, “Peopling of the New World.”
[57] Kemp, “Holocene Skeletal Remains.”
[58] Ugo A. Perego and others, “The Initial Peopling of the Americas: A Growing Number of Founding Mitochondrial Genomes from Beringia,”Genome Research20 (2010): 1174–79.
[59] Miroslava Derenko and others, “Origin and Post-Glacial Dispersal of Mitochondrial DNA Haplogroups C and D in Northern Asia,” PLoS One 5 (2010): e15214.
[60] Sigríður Sunna Ebenesaersdóttir and others, “A New Subclade of MtDNA Haplogroup C1 Found in Icelanders: Evidence of Pre-Columbian Contact?” American Journal of Physical Anthropology 144 (2011): 92–99.
[61] Perego, “The Initial Peopling.”
[62] Ripan S. Malhi and others, “Brief Communication: Mitochondrial Haplotype C4c Confirmed As a Founding Genome in the Americas,”American Journal of Physical Anthropology141 (2010): 494–97.
[63] Stanford and Bradley, “Ocean Trails and Prairie Paths?”; and Bradley and Stanford, “The North Atlantic ice-edge corridor.”
[64] Ebenesaersdóttir, “A New Subclade of MtDNA Haplogroup C1.”
[65] Perego, “Distinctive Paleo-Indian Migrations.”
[66] Ebenesaersdóttir, “A New Subclade of MtDNA Haplogroup C1.”
[67] Alessandro Achilli and others, “The Mitochondrial DNA Landscape of Modern Mexico,” American Society of Human Genetics, 58th Annual Meeting, Philadelphia, November 11–15, 2008; and Alessandro Achilli and others, “Decrypting the MtDNA Gene Pool of Modern Panamanians,” American Society of Human Genetics, 59th Annual Meeting, Honolulu, October 20–24, 2009. Approximately 80 percent of the samples tested for the Mexican (n = approx. 2,000) and the Panamanian (n = approx. 500) mixed populations belonged to one of the four Pan-American haplogroups.
[68] Kari B. Schroeder and others, “Haplotypic Background of a Private Allele at High Frequency in the Americas,” Molecular Biology and Evolution26 (2009): 995-1016.
[69] “A Quantum Leap in DNA Studies,” www.signaturebooks.com/
[70] Morten Rasmussen and others, “Ancient Human Genome Sequence of an Extinct Palaeo-Eskimo,”Nature463 (2010): 757–62.
[71] Cassandra Brooks, “First Ancient Human Sequenced,” www.thescientist.com/
[72] Cassandra Brooks, “First Ancient Human Sequenced.” The second quotation is Brooks’s paraphrase of Feldman. See Michael H. Crawford,The Origins of Native Americans, 4. In his lengthy review of data supporting the Asian origins of the Amerindians, he stated that “this evidence does not preclude the possibility of some small-scale cultural contacts between specific Amerindian societies and Asian or Oceanic seafarers.”
[73] Antonio Arnaiz-Villena and others, “The Origin of Amerindians and the Peopling of the Americas According to HLA Genes: Admixture with Asian and Pacific People,”Current Genomics11, no. 2 (2010): 103–14.
[74] Richard E. Green and others, “A Draft Sequence of the Neandertal Genome,” Science 328 (2010): 710–22.
[75] Signature Books, “Neanderthals and Lamanites: New Science Impact Book of Mormon DNA Studies,” http://
[76] Critics of the Book of Mormon regard this statement as a typical Mormon apologists’ favorite cop out to justify the lack of evidence of Israelite DNA in the Americas. In 2010, this argument was brought up again in a forum discussing Mesoamericans and their DNA lineages. One of the contributors wrote that the “absence of evidence IS evidence of absence if one could reasonably expect there to be evidence where there is none.” Richard Packham, “Absence of evidence is not evidence of…”?? Posted on August 15, 2010 at http://
[77] See, for example, the introduction to Southerton,Losing a Lost Tribe (Sourceflix, 2003), DVD;or Living Hope Ministries,DNA vs. the Book of Mormon(Sourceflix, 2003), DVD.
[78] “What Happens Genetically When a Small Population Is Introduced into a Larger One?” www.signaturebooks.com/
[79] Dennis H. O’Rourke and Jennifer A. Raff, “The Human Genetic History of the Americas: The Final Frontier,”Current Biology20, no. 4 (2010): R202–7.
[80] Ugo A. Perego, “Current Biology, SMGF, and Lamanites,” www.fairblog.org/
[81] Jeffrey R. Holland, “Safety for the Soul,”Ensign, November 2009, 88–90, http://
